![]() Número 26,
julio, 2025: 1–10
Número 26,
julio, 2025: 1–10
ISSN versión impresa: 2071–9841 ISSN versión en línea: 2079–0139 https://doi.org/10.33800/nc.vi26.374
A NEW SPECIES OF CREMATOGASTER
(HYMENOPTERA: FORMICIDAE: MYRMICINAE)
WITH CONTRASTING COLORATION UNIQUE TO THE CARIBBEAN
Una nueva especie de Crematogaster (Hymenoptera: Formicidae: Myrmicinae) con coloración contrastante única del Caribe
Gianpiero Fiorentino1*, América Sánchez2 and Phillip Barden 1a3
1Federated Department of
Biological Sciences, New Jersey Institute of Technology, New Jersey, USA. abarden@njit.edu, ![]() https://orcid.org/0000-0001-6277-320X.
https://orcid.org/0000-0001-6277-320X.
2Departamento de Investigación y
Conservación, Museo Nacional de Historia Natural “Prof. Eugenio de Jesús
Marcano”, Santo Domingo, Dominican Republic. a.sanchez@mnhn.gov.do, ![]() https://orcid.org/0000-0002-1540-8493.
https://orcid.org/0000-0002-1540-8493.
3Division of Invertebrate
Zoology, American Museum of Natural History, New York, USA. *Corresponding
author: gf6@njit.edu, ![]() https://orcid.org/0000-0001-6948-5032.
https://orcid.org/0000-0001-6948-5032.
[Received: January 9, 2025. Accepted: June 20, 2025]
ABSTRACT
The cosmopolitan ant genus Crematogaster is a conspicuous lineage of myrmicine ants. Here, we describe a new species from Hispaniola, Crematogaster dorada sp. nov. C. dorada is readily diagnosable from other Caribbean ants through mesosomal thoracic sculpturing and a unique color pattern; its taxonomic affinities are consistent with other members of the Neocrema clade in the Orthocrema group, a lineage with no previous record in the Caribbean. The current diversity of Greater Antilles Crematogaster now stands at 10 species and three putative subspecies. Expanded sampling will likely yield additional species as much of the Caribbean fauna has been unrevised since the early 20th century.
Keywords: Crematogaster, Caribbean, Greater Antilles, color morph.
RESUMEN
El género Crematogaster es un linaje conspicuo de hormigas mirmicinas distribuidas globalmente. Aquí, describimos una nueva especie para la isla Hispaniola, Crematogaster dorada sp. nov. C. dorada se diferencia fácilmente de otras hormigas del Caribe por la escultura torácica del mesosoma y un patrón de color único; sus afinidades taxonómicas son consistentes con miembros del clado Neocrema, un linaje sin registro previo en el Caribe. La diversidad actual de Crematogaster en las Antillas Mayores asciende a 10 especies y tres subespecies putativas. Es probable que un muestreo expandido revele especies adicionales, dado que gran parte de la fauna del Caribe no ha sido revisada desde principios del siglo XX.
Palabras clave: Crematogaster, Caribe, Antillas Mayores, morfotipo de color
INTRODUCTION
The genus Crematogaster is a globally distributed lineage of Myrmicinae ants with a distinct morphological habitus. Workers in this group possess an anteromedially-compressed gaster that resembles a heart shape (Bolton, 2003). This abdominal conformation allows workers to raise the gaster over the mesosoma in a defensive posture while venom remains exuded from sting lancets (Kugler, 1979). Species richness for the group is greatest in the Afrotropics; however, species are documented from Canada to Argentina in the Western Hemisphere; some species are well-distributed members of Neotropical ecosystems in particular. Most neotropical diversity resides within the Orthocrema-clade and is the result of two dispersals from the Afrotropics and Southeast Asia during the Oligocene-Miocene (Blaimer, 2012a).
While there are dozens of described Central and South American species, the Greater Antilles harbors just 10 known Crematogaster species (including a species newly reported here). Here, we describe a new species of Crematogaster from Hispaniola, which represents just the third known from the island.
OBJECTIVES
- To describe a new species of Crematogaster from the southeast of the island of Hispaniola.
MATERIALS AND METHODS
Imaging and morphological terminology
The examined specimens are deposited in the Museo Nacional de Historia Natural “Prof. Eugenio de Jesús Marcano” located in Santo Domingo, Dominican Republic. For the preliminary placement of the studied specimens, we used the worker-based key to Crematogaster species proposed by Longino (2003). The morphological terminology follows Bolton (1994), for most body structures, and Longino (2003) and Blaimer (2012b) for Crematogaster-specific terminology. For measurements, we followed those taken for Costa Rican Crematogaster ants following Longino (2003). Specimens were observed using a Nikon SMZ25 stereomicroscope. Measurements were taken using a micrometer lens at magnifications of 60-80x. All measurements (Fig. 1, Table S1) are presented in mm.
Color morph diversity
To assess the variation across Caribbean Crematogaster species, we compiled color values from specimens and species descriptions. Specimen-based colors were obtained from AntWeb and photos produced for the description herein, both sources were based on dorsal stereoscope images. For stereoscope images, we obtained average RGB values for segments by taking an average of pixel color values from an unoccluded (i.e. no debris or light reflection) region of both the head and gaster. We could not obtain specimens or AntWeb images for a subset of taxa, in these cases we generated RGB estimates from primary species descriptions. For example, the description of C. manni (Buren, 1968) provides a color of “dark brown” for the head and “black” for the gaster, yielding approximate RGB values of 53, 33, 0 and 0, 0, 0. The accuracy of this sampling scheme is low based on the subjective nature of qualitative color text descriptions and camera variation for specimen-based sampling, however, the relative contrast between head and gaster color is of primary interest given the unique morphology of the taxon described here. This broad-level intra-specimen contrast can be considered reliable and is apparent in our diversity sampling. All sampling sources and RGB values are provided in Supplemental Dataset 1.
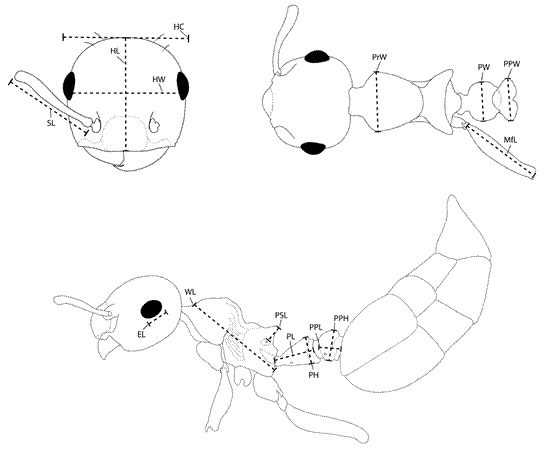
Figure 1. Glossary of morphological terminology used. HL, head length; HW, head width; HC, head capsule width; SL, scape length; EL, eye length; A11L, A10L, A09L, A08L, length of 11th to 8th antennal segment; A11W, A10W, A09W, A08W, width of 11th to 8th antennal segment; WL, Weber’s length; SPL, propodeal spine length; PTH, petiole height; PTL, petiolar length; PTW, petiolar width; PPL, post Petiolar length; PPW, post petiolar width; CI, cephalic index; OI, ocular index; SI, scape index.
RESULTS
Systematics
Order Hymenoptera Linnaeus, 1758
Family Formicidae Latreille, 1809
Subfamily Myrmicinae Lepeletier de Saint-Fargeau, 1835 Genus Crematogaster Lund, 1831 Crematoagster dorada sp. nov.
urn:lsid:zoobank.org:pub:EB52AF2D-B400-4697-98AC-71AAAFEDC3CB
Figs. 1–4

Figure 2. Photomicrographs of Crematogaster dorada sp. nov. worker holotype MNHNSD 18.112. A, head in full-face view; B, dorsal view; C, lateral view. Scale bar = 0.5 mm.
Type material
Holotype worker, Provincia Santiago, San José de las Matas, centro El Pinar, 10-IV2014, G. de los Santos, 19º22’24.1’’ N 071º01’41.4’’ W, 559 msnm, deposited in MNHNSD under catalog number MNHNSD 18.112. Paratypes, same data as holotype, deposited in MNHNSD under catalog numbers: MNHNSD 18.110, MNHNSD 18.111, MNHNSD 18.115, MNHNSD 18.116, and the Museum of Comparative Zoology (MCZ) under catalogue numbers:
MCZ-ENT00828245 and MCZ-ENT00828246.
Diagnosis (English)
Color dark red-brown to black with strikingly yellow golden gaster, head in lateral view with bulbous clypeus, head in face view with two rows of erect setae and 2–3 erect setae on vertexal sides of head; pronotum short and strongly convex, posterior face of mesonotum with a straight declining portion followed a second declining face dropping steeply to propodeal suture, propodeum strongly swollen, with completely blunt and rounded propodeal spines; propodeal spiracle conspicuous and oval; petiole wedge-shaped, ovo-rectangular and wider than long in dorsal view, ventral tooth absent; postpetiole bilobed posteriorly, ventral tooth absent.
Diagnosis (español)
Color rojizo-marrón oscuro a negro con un gaster amarillo-dorado, cabeza en vista lateral con clípeo globoso, cabeza en vista frontal con dos hileras de pelos erectos y 2 a 3 pelos erectos en vértice de la cabeza; pronoto corto y fuertemente convexo, cara posterior del mesosoma con un ligero declive, seguido por una cara fuertemente angulada, bajando a la sutura propodeal; propodeo fuertemente hinchado, espinas propodeales romas y redondeadas, espiráculo porpodeal conspicuo y ovulado; peciolo en vista lateral, en forma de cuña, ovo-rectangular y más ancho que largo en vista dorsal, diente ventral ausente; pospeciolo bilobulado, diente ventral ausente.
Measurements - HL 0.77; HW 0.78; HC 0.83; SL .60; EL 0.14; A11L .27; A11W 0.12; A10L
0.16; A10W 0.10; A09L 0.09; A09W 0.07; A08L 0.06; A08W 0.07; WL 0.65; SPL 0.10; PTH
0.16; PTL 0.26; PTW 0.20; PPL 0.16; PPW 0.21; CI 101; OI 18; SI 78; PTWI77; PPI 131; ACI
1.04
Description
Worker - Color dark red-brown to black. Head sub-rounded, longer than wide (Fig. 2A); mandible largely smooth and shiny; clypeus smooth and shiny, straight anteriorly, bulbous, forming a notable protuberance in profile view; face smooth and shiny; scape with moderately abundant, short, appressed pilosity and no erect setae; terminal 2–3 segments of antenna sequentially longer and more densely pubescent, especially terminal two, forming two-segmented club; face and ventral surface of head with sparse short appressed pubescence, face with six erect setae organized in two rows of three; 2–3 erect setae projecting from vertexal sides of head in full-face view.
In lateral view, promesonotum compressed, pronotum short but strongly convex, mesonotum projecting above pronotum and propodeum (Fig. 2C); posterior face of mesonotum with a straight declining portion followed a second declining face dropping steeply to propodeal suture; mesopleuron mostly smooth and shiny; katepisternum and anapisternum not separated by suture, both strongly sculptured; humeral corners with a pair of setae; mesonotum with two pairs of setae, one long and one short; propodeum with a short anterior bulge, followed by a straight an flat face leading into the propodeal spines; propodeal suture deep, V-shaped, but suture partially obscured in lateral view by small lateral carinulae that bridge suture (Fig. 2B); propodeum strongly swollen, with completely blunt and rounded propodeal spines (Fig. 3B); propodeal spiracle conspicuous and oval; in dorsal view, pronotum slightly angled and smooth, mesonotum is strongly striated, with longitudinal striations that curve over the mesopleuron; mesometanotal suture strongly marked; promesonotal suture not well developed, but segments are easily distinguishable by their sculpturing; the propodeum is swollen, and propodeal spines are swollen as well, the space between the spines is smooth and shiny; legs with sparse, fully appressed pubescence.
Petiole in side view wedge-shaped, mostly smooth with faint striated surface sculpture; anteroventral tooth absent; dorsal face subrectangular, slightly wider than long, smooth and shiny, anterolateral portions somewhat produced as lobes, anteromedian region depressed, sides rounded anteriorly, converging posteriorly (Fig. 3A); postpetiole ventral tooth absent, postpetiole in dorsal view bilobed, wider than long, with deep median longitudinal impression, posterior margin emarginate; fourth abdominal tergite smooth or with faint microareolate sculpture; pair of stiff setae on posterodorsal petiole, pair on postpetiole on each lobe, and more than 20 stiff setae on fourth abdominal tergite.
Etymology
The name “dorada” is derived from the Spanish word for golden and refers to the species’ distinct golden gaster.
Comments
Crematogaster dorada sp. nov. belongs to the Orthocrema sub-group, given the distinctly bilobed postpetiole with deep median longitudinal impression, an ovo-retangular petiole with dorsoposterior lateral tubercules, each bearing an erect seta, and a rounded posterior head margin (Blaimer, 2012b). C. dorada also represents the first record of Crematogaster (Orthocrema) of the Neocrema clade for the Caribbean and may be of significance towards understanding the history of the genus in the region (Ward & Blaimer, 2022). This species stands out from all other Crematogaster for its striking coloration, workers are distinctly bi-colored with a dark brown head, mesosoma and legs, and a golden orange gaster. Crematogaster dorada is highly autapomorphic, possessing the combination of a bulbous clypeus, completely blunt and rounded propodeal spines, and the distinct lack of petiolar or postpetiolar anteroventral teeth. Though the species possesses some similarity in propodeal spine configuration to C. curvispinosa (broad divergent bases), the spines are never the same as in C. dorada (blunt and rounded); they more closely resemble C. raptor, although only in the bluntness of the spines. Crematogaster dorada is a distinct taxon that, although referenced by Lubertazzi (2019) and Prebus (2021), had yet to be formally described.

Figure 3. Enlarged stereoscope images of C. dorada holotype MNHNSD 18.112. A, dorsal view of head, mesosoma, and petiole. B, lateral view of mesosoma. Scale bar = 0.25 mm
DISCUSSION
Despite the low diversity of Crematogaster in the Caribbean, with just 10 described species, Crematogaster dorada stands out for its bi-colored body (Fig. 4). Yet, this bi-color pattern is shared by other distantly-related island endemics, such as Cephalotes unimaculatus (Smith), Linepithema keiteli (Forel), Nylanderia disatra (LaPolla & Kallal), Pheidole harlequina (Wilson), and Temnothorax harlequina (Prebus), and may be common in Mesoamerican and Greater Antillean ant fauna (Prebus, 2021). Among the Crematogaster of the Caribbean, however, this color morph is singular (Fig. 4).
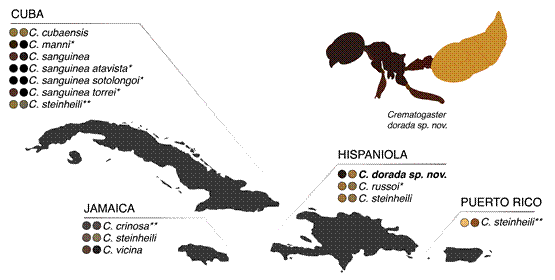
Figure 4. Distribution of Greater Antilles Crematogaster species and subspecies by color morph. For each species, the left circle denotes the color of head, the right circle denotes the gaster color. Colors extracted from primary and AntWeb stereoscope images as well as qualitative primary literature descriptions. Taxa with one asterisk (*) denote colors extracted from RGB estimates of qualitative primary literature descriptions, two asterisks (**) indicate a stereoscope sample but from a location other than the island of interest (e.g. we could not obtain images of C. steinheili sampled in Cuba and so include colors based on a specimen from Jamaica). A full dataset of sampled specimens, color sources, and RGB values is available in Supplemental Dataset 1.
This pattern is most prevalent among Pheidole, Temnothorax, and Cephalotes species on the island (Fig. 5), which may indicate a potential case of color mimicry within these genera (Lubertazzi, 2019; Prebus, 2021). While this coloration is not found in other Caribbean Crematogaster, this may be due to limited sampling of Haiti, as opposed to the island’s eastern side of the Dominican Republic (Lubertazzi, 2019). The color mimicry found particularly in small-bodied ants in the Dominican Republic could be evidence of a Müllerian mimicry complex, a Batesian mimicry complex, or broadly applicable environmental selective pressure. Color mimicry complexes, though known from other Hymenoptera (Wilson et al., 2015; Wilson et al., 2022), have not been reported in ants.
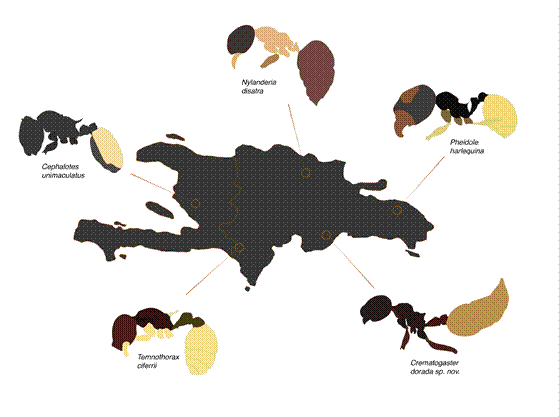
Figure 5. Distribution of Hispaniolan ants with similar bicolored color patterns. The five species highlighted here correspond to five distinct genera that have independently developed similar color morphs defined by contrasting colors between the gaster and the rest of the body. Arrows indicate the approximate known distribution of the ant species.
ACKNOWLEDGMENTS
This material is based upon work supported by the National Science Foundation under Grant No. 2144915 to Barden and the Theodore Roosevelt Memorial Fund of the American Museum of Natural History.
REFERENCES
Blaimer, B. B. (2012a). Acrobat ants go global: Origin, evolution, and systematics of the genus Crematogaster (Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution, 65(2), 421–436.
Blaimer, B. B. (2012b). A subgeneric revision of Crematogaster and discussion of regional species-groups (Hymenoptera: Formicidae). Zootaxa, 3482, 47–67.
Bolton, B. (1994). Identification guide to the ant genera of the world. Cambridge, Mass.: Harvard University Press, 222 pp.
Bolton, B. (2003). Synopsis and classification of Formicidae. Memoirs of the American Entomological Institute, 71, 1–370.
Buren, W. F. (1968). A review of the species of Crematogaster, sensu stricto, in North America (Hymenoptera, Formicidae). Part II. Descriptions of new species. Journal of the Georgia Entomological Society, 3, 91–121.
Kugler, C. (1979). Evolution of the sting apparatus in the myrmicine ants. Evolution, 33(1), 117–130.
Longino, J. T. (2003). The Crematogaster (Hymenoptera, Formicidae, Myrmicinae) of Costa Rica. Zootaxa, 151, 1–150.
Lubertazzi, D. (2019). The ants of Hispaniola. Bulletin of the Museum of Comparative Zoology, 162(2), 59–210.
Prebus, M. M. (2021). Taxonomic revision of the Temnothorax salvini clade (Hymenoptera: Formicidae), with a key to the clades of New World Temnothorax. PeerJ, 9, e11514. https://doi.org/10.7717/peerj.11514
Ward, P.S., Blaimer, B. (2022). Taxonomy in the phylogenomic era: species boundaries and phylogenetic relationships among North American ants of the Crematogaster scutellaris group (Formicidae: Hymenoptera). Zoological Journal of the Linnean Society, 194(3), 893–937. https://doi.org/10.1093/zoolinnean/zlab047
Wilson, J. S., Jahner, J. P., Forister, M. L., Sheehan, E. S., Williams, K. A., & Pitts, J. P. (2015). North American velvet ants form one of the world’s largest known Müllerian mimicry complexes. Current Biology, 25(15), R704–R706.
Wilson, J. S., Pan, A. D., Alvarez, S. I., & Carril, O. M. (2022). Assessing Müllerian mimicry in North American bumble bees using human perception. Scientific Reports, 12(1), 17604. https://doi.org/10.1038/s41598-022-22402-x
Suppplementary
Table S1. Glossary of morphological measurements taken from the specimens. Modified from Longino (2003).
|
HL |
Head length. In full-face view, the maximum distance from the posterior margin of the head to the anterior margin of the clypeus. |
|
HW |
Head width. In full-face view, the maximum width of the head, excluding the eyes. |
|
HC |
Head capsule width. maximum width of head in full-face view, excluding eyes; may be anterior or posterior to eyes. |
|
SL |
Scape length. In frontal view, the maximum length of the scape excluding the basal condyle and neck. |
|
EL |
Eye length. In lateral view, eye length is measured along its maximum length. |
|
A11L, A10L, A09L, A08L |
Length of 11th to 8th antennal segment. |
|
A11W, A10W, A09W, A08W |
Width of 11th to 8th antennal segment. |
|
WL |
Weber’s length. In lateral view, the distance between the anterior margin of the pronotum, excluding the collar, to the posteroventral margin of the metapleuron. |
|
SPL |
Propodeal spine length. Measured from tip of propodeal spine to closest point on outer rim of propodeal spiracle. |
|
PTH |
Petiole height. Viewed in lateral profile, perpendicular distance from ventral margin to highest point of posterolateral tubercles; if ventral margin is concave upward, measured from line tangent to uppermost portion of curve and oriented as close as possible to long axis of petiole. |
|
PTL |
Petiolar length. In lateral view, the distance between the anterior margin of the petiole, including its anterolateral projection, to its posterior margin, excluding the posteroventral folded ridge that embraces the helcium. |
|
PTW |
Petiolar width. In dorsal view, the maximum width of the petiole. |
|
PPL |
Post-petiolar length. In lateral view, the distance between the anterior margin of the petiole, including its anterolateral projection, to its posterior margin, excluding the posteroventral folded ridge that embraces the helcium. |
|
PPW |
Post-petiolar width. In dorsal view, the maximum width of the petiole. |
|
CI |
Cephalic index; 100*HW/HL. |
|
OI |
Ocular index; 100*EL/HL. |
|
SI |
Scape index; 100*SL/HL. |
Citation: Fiorentino, G., Sánchez, A., & Barden, P. (2025). A new species of Crematogaster (Hymenoptera: Formicidae: Myrmicinae) with contrasting coloration unique to the Caribbean.
Novitates Caribaea, (26), 1–10. https://doi.org/10.33800/nc.vi26.374